|
|
|
 |
|
|
Sherrington called the motor neuron the final common pathway. All the subtle signals converging from several descending tracts as well as afferent input from the periphery are somehow integrated on the motor neuron, which subsequently conducts the appropriate signal out to the muscle. Because so many different pathways converge on the motor neuron, the contribution of any single tract to the final motor act is extremely difficult to determine. Several descending pathways have been shown to effect changes in the activity of motor neurons. The anatomical courses of these pathways have been extensively studied from their origins in various areas of the brain to their synaptic contacts with the motor neurons. The precise physiological roles of these pathways have been studied but the information is limited because of several factors. Principal among them is the fact that most of the work has concerned the motor neurons innervating hind limbs of the cat. Studies on primates have been continuing, but a big problem is the somewhat suspect attempt to wed the neurophysiology of the cat's movement performance to the neuroanatomy of the human's. Another problem lies in the fact that a common tool for studying the function of nerve pathways is electrical stimulation. While there seems to be little alternative to this procedure, the meaningfulness of artificially induced volleys of impulses is questionable when one considers that tile natural influences on motor neurons are spatially and temporally varied and probably achieve their effects by virtue of a pattern of impulses rather than a repetitive volley. Recent attempts have been made to study the neurophysiology of movement by recording neuromuscular potentials accompanying spontaneous movement. This is certainly a desirable approach but is also limited by the fact that even simple body movements are neurally very complex. Thus attempts to relate the anatomical and physiological events associated with these movements are difficult and hard to interpret. Nevertheless, much has been learned concerning the role of the nervous system in such activities as walking, running, and the regulation of postural movements. It now appears that there are "pattern generators" or "prewired" groups of neurons within the central nervous system producing a wide variety of basic motor programs. "Command" neurons activate these pattern generators when a particular movement is called for. Here, we will examine some of these pattern generators as well as the role of the brain and its descending pathways in initiating and regulating movement.
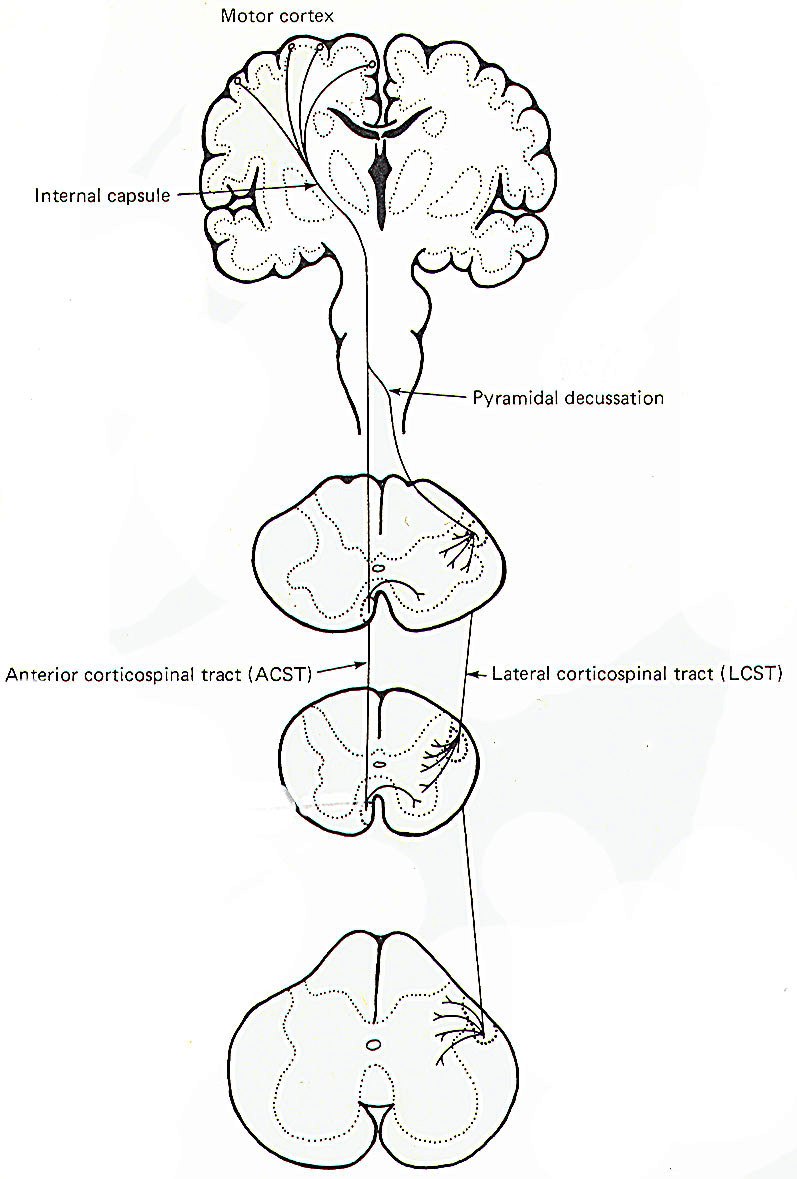
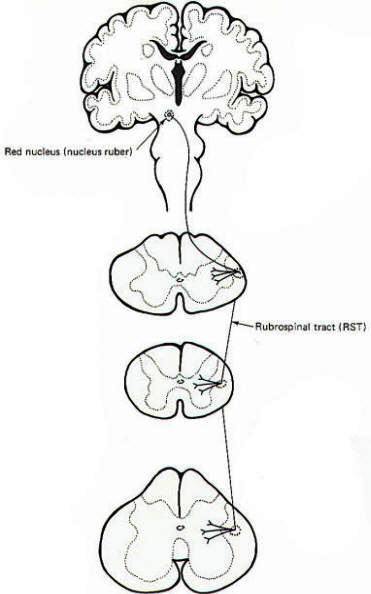
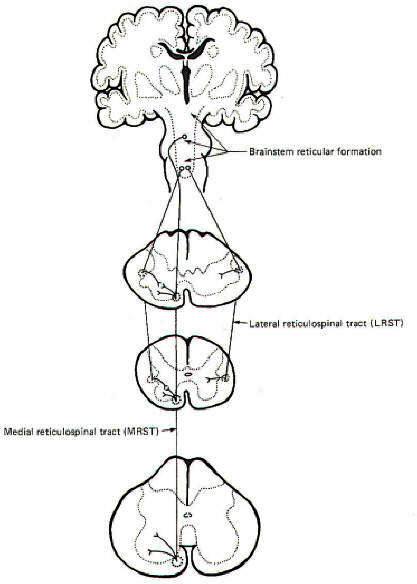
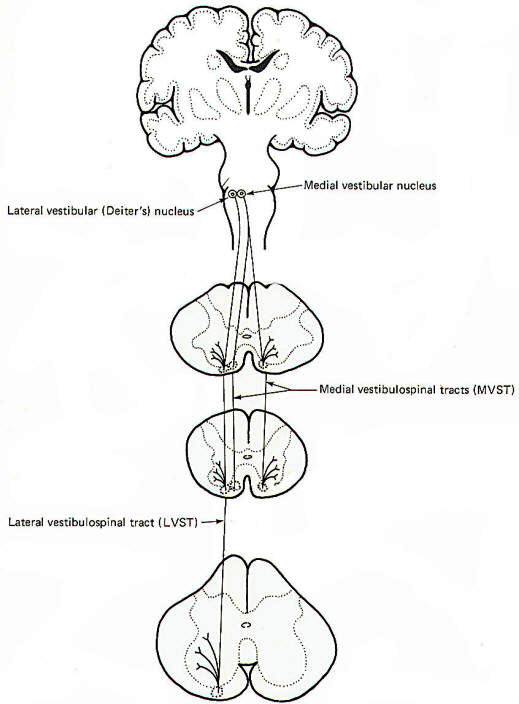
Electrophysiological studies have shown that the motor cortex resembles a map showing a distorted image of the body turned upside down and reversed left to right. Some motor pathways to the skeletal musculature of the body arise directly from cells within the cerebral motor cortex, while others arise from subcortical areas of the brain and brainstem. Neurons that originate in the cerebral motor cortex. the cerebellum, or various brainstem nuclei that send axon, into the brain stem and spinal cord to activate cranial or spinal motor neurons are called upper motor neurons, Those cranial and spinal motor neurons which actually innervate muscles are the lower motor neurons. The latter include the alpha and gamma motor neurons of spinal nerves. Upper motor neurons are found entirely within the CNS, while the fibers of lower motor neurons are part of the PNS. Upper motor neurons are clustered together to form descending tracts in the brain and spinal cord. Such tracts are commonly named according to their site of origin and the region of their distribution. An example is the corticospinal tract, which originates in the cerebral cortex and is distributed to the spinal cord. Another is the rubrospinal tract. which originates in the red nucleus (nucleus ruber) of the midbrain and is distributed to the spinal cord. The lower motor neurons of spinal nerves are somatotopically organized in the anterior horn of the spinal cord gray matter. In general, those innervating the distal limb musculature are located in the lateral aspects of the anterior horn, while those innervating proximal limb muscles are found in the intermediate region, The most medial group of motor neurons innervates the musculature of the appendicular and pelvic girdles.
Upper motor neurons don't simply stimulate lower motor neurons and produce movement. The highly skilled and coordinated movements of which humans are capable would seem to require a more complex and involved system. While little is known of the highly involved and integrated activity which occurs in the brain's neural circuits during even a simple body movement, it now appears that highly coordinated and very complex systems of interneurons regulate the precise timing and sequencing of muscle activity which is observed in such movements. There is also increasing evidence that groups of interneurons cause specific patterns of impulses to fire in the lower motor neurons associated with a given coordinated movement. The central theory is that these interneurons form pattern generators within the CNS which produce the basic motor program. At the spinal cord level the pattern generator is composed of a set of local control centers located in the gray matter. There are neurons within these centers which coordinate muscular synergies and generate timing signals. Command neurons activate these pattern generators when a particular coordinated movement is required. The result of such activation is that the lower motor neurons fire in a properly sequenced and timed pattern to produce a coordinated movement. Identification of specific command neurons for a particular human movement is a difficult process and a speculative one at best. It may be that upper motor neurons from the brain and brainstem function in this respect for voluntary movements and reflex postural adjustments. In some invertebrates, however, the activation of a single interneuron is sufficient to excite an entire coordinated muscle behavior. For example, stimulation of the giant axon of the crayfish produces a coordinated tail flip which propels it away from the stimulus. Similarly, stimulation of the Maunther cells of teleost fishes produces a tail flip propelling the fish away from the stimulus. These cells are part of the reticulospinal tract neurons in the fish and apparently serve as command neurons that activate the pattern generator which carries the motor program for tail flip. It is presumptuous to assume that all movements proceed in accordance with pattern generators and prewired motor programs. Nevertheless, it may be that certain basic coordinated movements of the limbs and trunk may proceed in a very general way under the influence of such programs, while the initiation and fine tuning of the movement requires input from descending and sensory pathways. The upper motor neurons of some descending motor pathways no doubt serve as command neurons for certain movement patterns. Variations in the discharge patterns of these neurons determines the variability of the programmed response. There is evidence that a change in the firing frequency of certain command neurons leads to a change in the intensity of the response. If the coordinated movement involves a postural system, altering the firing rate alters the magnitude of postural adjustment. If a locomotor system is involved, the frequency of the movement cycle will vary with changes in the frequency of command neuron firing. Other command neurons produce the same motor pattern regardless of their firing rates. Their role seems to be simply turning the program on and off. Still others may regulate the magnitude of the programmed response. Most of the vertebrate work involving motor programs has dealt with locomotor activity in the cat. Perhaps we can get a feel for the intricate features of such programs by an examination of this work.
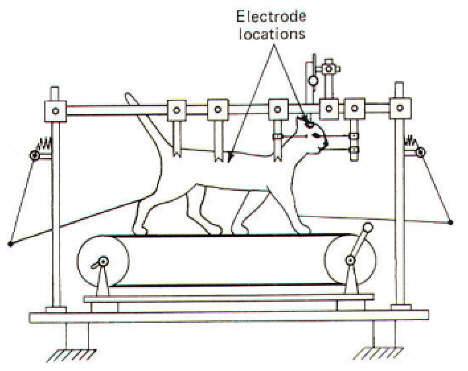
Removal of the telencephalon (cerebral hemispheres) and the rostral portion of the thalamus in an acute cat preparation remarkably leaves the animal with practically normal locomotion. It can walk spontaneously on its own and it can be forced to run by electrical stimulation of a region in the subthalamus called the subthalamic locomotor region (SLR). However, spontaneous walking movements cease in acute preparations where the brainstem is sectioned caudal to the subthalamus but just rostral to the midbrain. This is a mesencephalic preparation, meaning the highest intact brain component is the midbrain or mesencephalon. There are several advantages to the study of locomotion in such a preparation. Perhaps most important is that relatively normal locomotor movements can be initiated by the electrical stimulation of an area in the tectum of the midbrain called the mesencephalic locomotor region (MLR). Such a cat preparation can also be placed on a treadmill to facilitate natural movements while its head is fixed in a stereotaxic apparatus enabling the experimenter to conveniently stimulate various brain stem areas and observe the results (Fig-1).
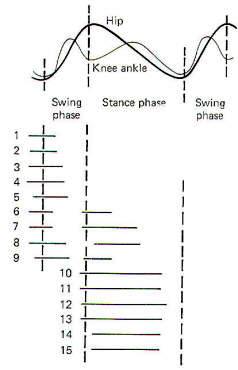
Walking and running movements in a mesencephalic cat are similar to those observed normally. A single step cycle is accomplished when a limb touches down, lifts off and moves forward, and then touches down again. The step cycle is composed of a stance phase (limb in contact with the ground) and a swing phase (limb lifted off and moving forward). Muscle group activity during the step cycle proceeds in a logical order. At the end of the stance phase, when the limb is at its most caudal position, the flexors become active, lifting the leg and initiating the swing phase. During the swing phase, the relaxation of the flexors combined with onset of extensor activity and inertia all propel the limb forward. Because the extensors begin to contract prior to the stance phase, the limb is able to support the weight of the body as the limb touches down. Extensor activity continues throughout the stance phase until just at the end, when it begins to diminish and is replaced by increasing flexor activity preceding another step cycle (Fig-2). Increasing the stimulation of the MLR causes an increase in the stepping frequency of the mesencephalic cat. However, this increased frequency is apparently due to an increased muscular force moving the treadmill faster with the rate of stepping increasing indirectly to keep up with it. The increased force is apparently due to the increased recruitment of more alpha motor neurons and motor units rather than to any increase in the firing rates of the currently active units. Thus MLR stimulation directly increases the level of muscular force and indirectly the stepping frequency. If the level of MLR stimulation is held constant and the treadmill is either speeded up or slowed down by the experimenter, the stepping frequency of the mesencephalic cat will speed up or slow down accordingly. It is very interesting to note that while stepping is a complex process involving a repetitive sequence of muscular contractions and relaxations with very precise timing, all that is necessary to get it started is to stimulate the MLR in the mesencephalic cat or the SLR in the subthalamic cat. Thus it seems likely that stepping is an automatic process with a central program controlled by a pattern generator in the CNS. Stimulation of the MLR and SLR activates this program and can in fact vary its intensity. The pattern generator for stepping with the hind limbs of the cat appears to reside in the spinal cord. In chronic cat preparations where the lower thoracic spinal cord was completely sectioned shortly after birth. the animals are capable of a full variety of stepping gaits in accordance with the speed of the treadmill. Thus it seems likely that the pattern generator resides in the spinal cord, at least for hind limb movements. Transection of the spinal cord at a high enough level to include the forelimbs (high cervical) does not ordinarily allow for satisfactory locomotor movements. and thus has not been adequately evaluated in this regard.
Signals arrive at the pattern generator in the spinal cord both from the periphery and from supraspinal levels. The Ia afferents from muscle spindles monosynaptically stimulate homonymous alpha motor neurons and thus influence the activity of an ongoing motor program. Similarly, signals arriving at spinal cord interneurons from supraspinal levels via upper motor neurons also exert an influence over the performance of a motor program. It is important to note that the stretch reflex is not always productively useful at all times during the step cycle. Thus it is not surprising to find that the sensitivity of the reflex is varied cyclically with the step. It is "tuned in" when the reflex is useful and "tuned out" when activation of the reflex would be counterproductive to a particular phase of the step cycle. Muscle spindle sensitivity can be controlled by the pattern generator since it can apparently direct the timing of both alpha and gamma motor neuron firing. Thus during the phase of the stepping cycle when a muscle is passively stretched (i.e., the gastrocnemius at the end of the stance phase) the sensitivity of its muscle spindles is decreased. This prevents the stretch reflex from activating muscle contraction during the "wrong" phase. Thus the spindles are "tuned out" when the muscle is passively stretched and "tuned in" again when the muscle becomes active during the step cycle. Let's now examine the descending motor pathways which influence motor activity.
|
|
|

Copyright [2007] [CNS Clinic-Jordan]. All rights reserved